
- Відвідувань: 6
- Переглядів: 6
Міф про древні ДНК-генеалогічні лінії африканців (частина 1)
Світ:
Спецтема:

Але розбиратися особливо і не треба, тому що у всіх чоловіків на Землі, африканців і неафриканців, у всякому разі, у всіх, хто робив тест на ДНК, сніп-ланцюжка Y-хромосоми по древності однакові. У всіх ці ланцюжки сніпів тягнуться від спільних предків з шимпанзе, орангутангом, горилою, макакою та іншими древніми спільними з нами предками. Нижче - характерний приклад, процитований ще в 2012 році в журналі Advances in Anthropology (Klyosov, Rozhanskii, Ryabchenko): 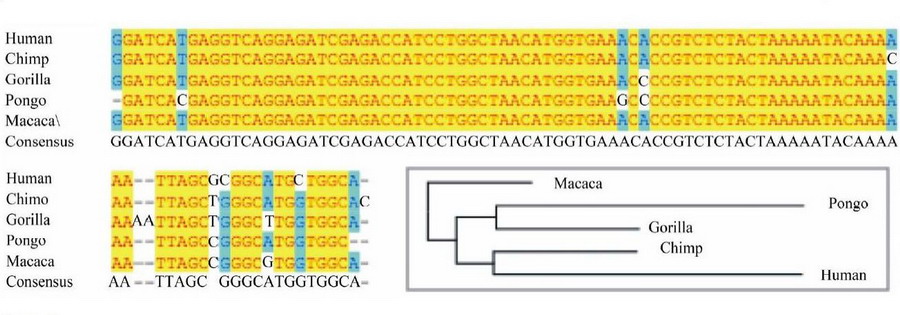
Порівняння фрагмента нуклеотидної послідовності Y-хромосоми
людини, шимпанзе, горили, орангутанга і макаки
Видно, що в 90% випадків нуклеотидні послідовності показаних фагментів Y-хромосоми збігаються. А у шимпанзе і людини збігаються 93 нуклеотиди з 97, тобто в 96%. Так що у всіх нас ДНК-коріння древні, і африканці в цьому відношенні ніяк не виділяються.
Добре, а наскільки стародавні? Антропологи оцінюють час розбіжності лінії майбутньої людини від лінії майбутнього шимпанзе в 4-6 мільйонів років тому, це ж повторюють і генетики, хоча пошуки статей з конкретними розрахунками результатів не дали. А ось результати розрахунку за допомогою автоматичного калькулятора Килина-Клёсова (ККК, 2015), ілюстрація збільшується по кліку: 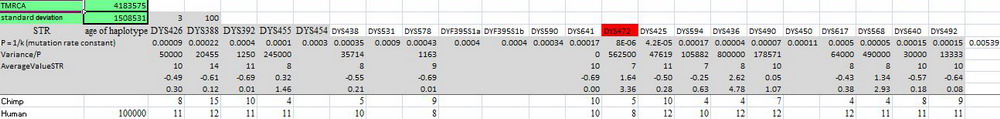
Спільний предок людини і шимпанзе жив, якщо округлити розрахунки, 4.2±1.5 мільйонів років тому. 16-маркерний гаплотип шимпанзе був витягнутий з геномної бази даних, як описано в тій статті в Advances in Anthropology (2012), а в якості узагальненого гаплотипу людини брався екстрапольований спадковий гаплотип по всіх неафриканських гаплогруп, перерахований на часи 100 тис. років тому. Втім, можна було і не перераховувати, ось - час до загального предка сучасних носіїв гаплогрупи А00 і шимпанзе, ілюстрація збільшується по кліку: 
Воно дорівнює 4.4±1.4 мільйонів років тому, що практично однаково в межах похибки розрахунків. Ті ж часи дають всі основні гаплогрупи серії А, наприклад:
• А0а-V150 має спільного предка з шимпанзе 4.6±1.7 млн років тому;
• A1b1-M32 (Чад і Туреччина) 4.3±1.4 млн років тому;
• A1b1-M32, M13 (Британія) 4.2±1.4 млн років тому, а також
• гаплогруппа В, 4.4±1.4 млн років тому.
Коротше, щодо «древніх ДНК-ліній» африканці нічим від неафриканців не відрізняються. І ті, й інші тягнуть свої ДНК-лінії від загального предка з шимпанзе. І в тих, і в інших ланцюжка сніпів йдуть на глибини в сотні тисяч і мільйони років. До цього ми ще повернемося.
Може, під «древніми африканськими ДНК-лініями» попгенетики та інші генетики натякають на стародавні часи життя їх спільних предків, і вважають, що вони набагато давніші, ніж у неафриканців? Доведеться їх засмутити, ситуація протилежна. Африканські предки групи А (популяцій, субкладів) майже всі зовсім недавні. Візьмемо, наприклад, гаплогруппу А00, яка серед гаплогрупп вважається найдавнішою. Так, для неї ланцюжок сніпів налічує - за даними, наведеними YFull - 1684 сніпів, що при 144 роках на сніп (це число призводить YFull) дає 242500 років від початку гаплогрупи. Дійсно, YFull призводять близький «вік» А00, 235000 років. Але звідки цей ланцюжок сніпів веде - ніхто не знає. Може, з Африки, може, з Кавказу, може, з Сибіру, може, з Європи - невідомо. Чомусь вважається, що саме з Африки. Але це знову «по понятіям», щоправда, звідки з'явилися, незрозуміло, вибачте за гру слів. Найдавніші ДНК знаходили в Європі, на Руській рівнині, в Сибіру, а в Африці жодного разу, правда, і досліджень таких не було. Так звідки ці «поняття» з'явилися? А так, стиль попгенетиків.
До речі, про вік гаплогрупи А00. Калькулятор КК дає для базових гаплотипів А0 і А00 вік загального предка (з округленням) 217000±87000 років. Це - вік А00 або трохи старше. Так що датування за СНіП відтворюється за гаплотипом в межах похибки розрахунків (ілюстрація нижче збільшується по кліку). 
Так от, про спільних предків африканських популяцій, субкладів групи А. Вони майже всі за часом дуже дрібні, часто всього кілька сотень років. Іноді - між 1000 і 2000 років. Найдавніші - приблизно 5000-7000 років. І знову - де ці предки жили, ніхто не знає. Погляньмо на гаплогрупи А00. У літературі (Mendez et al., 2013) описані 11 гаплотипів в 95-маркерному форматі (без 16 мультимаркерних гаплотипів) носіїв гаплогрупи А00, все з племені Mbo з Камеруну, і ще один - афроамериканець. Розрахунків в статті, звичайно, немає. Всі 11 гаплотипів містять 29 мутацій, що дає 29/11/0.161 = 16 умовних поколінь, але з урахуванням того, що 45 алелій (з тисячі сорок п'ять алелій) автори визначити не змогли, то до загального предка насправді 17 умовних поколінь, тобто 425±90 років.
Отже, сама гаплогруппа А00 утворилася більше 200 тисяч років тому, причому невідомо де, а загальний предок носіїв А00 з Камеруну жив приблизно 425 років тому. Якщо додати до серії з Камеруну ще гаплотип афроамериканця тієї ж гаплогрупи, який відрізняється від предкового гаплотипу камерунців на 11 мутацій, тобто на 11/0.161 = 68 → 73 умовних покоління, тобто на 1825 років, то їх загальний предок, камерунців і афроамериканця, жив приблизно (425+1825)/2 = 1125 років тому.
Прикладів недавніх спільних предків різних чоловічих популяцій гаплогруп класу А, які зараз живуть в Африці, можна навести чимало. У більшості цих популяцій гаплотипи або однакові або майже однакові, навіть для протяжних гаплотипів. З доступними гаплотипами гаплогрупи А00 ми вже розібралися, їм чотириста років. Погляньмо тепер на Проект FTDNA гаплогрупи А:
• У гаплогрупі/субкладі А0-V150 є два 25-маркерних гаплотипа, обидва ідентичні один одному, тобто їх загальний предок жив не більше 300-400 років тому. Інших гаплотипів в 25-маркерне форматі або вище немає.
• У гаплогрупі A0a-L981 (L994, V148, V166) між двома доступними 67-маркерними гаплотипами є 8 мутацій, що поміщає їх загального предка на 900 років тому.
• У гаплогрупі A0b-V166, L896 шостій 37-маркерних гаплотипів мають спільного предка 475±160 років тому.
• У гаплогрупі A1a-M31 («європейський кластер») п`ять 37-маркерних гаплотипів мають спільного предка 1 175±290 років тому.
• У гаплогрупі A1a-M31 (всі невідомого походження) п`ять 37-маркерних гаплотипів мають спільного предка 1880±390 років тому.
• У гаплогрупі A1a-M31 (з 18 гаплотипів - 14 невідомого походження, два з Англії, один з Швейцарії, один Кейп Верде) всі 37-маркерні, всі мають спільного предка 730±130 років тому.
• У гаплогрупі A1a-M31 («британський кластер») 19 гаплотипів в 67-маркерному форматі мають спільного предка 740±120 років тому.
У підсумку, 50 гаплотипів в 37-маркерному форматі чотирьох попередніх серій (А0а-М31) мають спільного предка 9270±970 років тому. Але треба звернути увагу, що це чотири серії гаплотипів, розкидані по різних континентах. Це - не одна «ДНК-лінія», це - чотири різних ДНК-лінії, з чотирма загальними предками, кожен «глибиною» від 700 до 1900 років.
Ну і де, дозволено запитати, «найдавніші ДНК-лінії в Африці»? Це - міф, нічим не обгрунтований. Копалин ДНК немає, древніх ліній, як ми бачимо, немає, до того ж, в Африці зовсім не гаплотипи групи А явяются домінуючими. Домінує гаплогруппа E1b; в Камеруні і Чаді, і почасти в Нігерії, домінує гаплогрупа R1b, тому не дивно, що в Африці спостерігається «висока генетична різноманітність», тільки це не від давнини, а від великої кількості носіїв різних гаплогруп, прибулих в Африку в різні часи. Та й взагалі, як можна серйозно звертати увагу на «генетичне різноманіття» у відкритих системах, якою є Африка? Як я вже не раз відзначав, самим «генетично різноманітним» містом світу є, напевно, Нью-Йорк, але навряд чи хтось буде серйозно стверджувати, що Нью-Йорк - найдавніше місто світу. Крім, напевно, популяційних генетиків, з їх примітивними і бездумними критеріями.
Звідки ж з'явився цей міф про «давнину африканських ліній»? Датувань адже їх до останнього часу не було, та й датування бувають, як ми переконалися, кількох видів - ланцюжків мутацій, що йдуть далеко вглиб, до приматів; ланцюжків мутацій, що ведуть до часів утворення відповідних гаплогруп; і датувань спільних предків сучасних носіїв відповідних гаплогрупп і субкладів. Жодна з цих датувань не "вкорінена» територіально, тобто ми не знаємо, на якій території, на якому континенті утворилася та чи інша гаплогрупа. Ми можемо тільки набирати відомості і складати фрагменти загальної картини, але Африка в цьому відношенні знаходиться на одному з останніх місць за накопиченим знанням. Але фантазій, натяжок і відвертих фальсифікацій про них з боку попгенетиків - хоч відбавляй.
Власне, це характерно для популяційних генетиків, але вони уроки з принципових помилок не витягають. Ще не так давно вони стверджували, що гаплогрупа R1b утворилася в Європі 30000 років тому, і спочатку прийняли в штики, а потім геть ігнорували відомості і датування ДНК-генеалогії, що R1b прибула з Південного Сибіру довгим міграційним шляхом до Європи приблизно 4 800 років тому (Klyosov, 2009; Klyosov, 2012). Називали це «псевдонаукою», особливо в цьому старалися попгенетики Балановське. Розворот стався в 2013 році, коли неподалік від Байкалу знайшли викопну гаплогрупу R з археологічною датуванням 24000 років тому. Крига зрушилася, і попгенетики, не моргнувши оком, не винних і не цитуючи статті до датувань ДНК-генеалогії, стали повторювати те, що вже було опубліковано нами кілька років тому. Наприклад, у недавній статті Batini, Jobling та інших (2015) датування гаплогрупи R1b в Європі вже даються як R1b-L11 4510 років (довірчий інтервал 3920-5160 років тому) і R1b-P312 4210 років (довірчий інтервал 3700-4790 років тому) . YFull дають датування R1b-U106 4900 років (довірчий інтервал 4400-5400 років тому), R1b-P312 4600 років (для TMRCA, довірчий інтервал 4300-5000 років тому).
На жаль, доводиться повторювати, що попгенетики уроків зі своїх помилок не витягають. Ситуація повторюється з Африкою. Даних немає, або дані говорять про інше, але їх придумують і спотворюють. Навіщо це їм потрібно? Для нормальної людини звучить неймовірно, але це ментальність васалів, які підлабузнюються перед сюзереном. Сюзерен вирішує, кого пригріти, кого запросити на конференцію, кому дати грант, кого взяти в колективну статтю, ввести в комісію або комітет, загалом, в інфраструктуру. Без сюзерена васал - ніхто, і васал це прекрасно розуміє. А навіщо це потрібно сюзерену? О, там велика політика. Васала до цих питань не допускають. Васал знає, що якщо він публічно засумнівається в африканське походження людства, то його дні полічені. Тому він, васал, готовий розірвати на частини будь-якого, хто сумнівається або виставляє дані проти. А оскільки він розірвати на частини не може, то мета - знайти компромат, сфабрикувати брехню, пересмикнути, спотворити - інших способів або аргументів у васала немає, у всякому разі, наукових. Подивимося на схему гаплогруп і субкладів групи А, наведену згідно класифікації ISOGG, в скороченому вигляді:

У цій схемі - джерело принципової плутанини попгенетиків. Вони не змогли побачити і усвідомити, що ця драбинка субкладів являє собою серію вилок, які періодично розходяться на «африканські» (за сьогоднішнім місцем проживання) і неафриканські гілки. Опишемо коротко ці вилки, а потім розглянемо їх докладніше - коли вони утворилися, і хто є нащадками субкладів по обидві сторони тих виделок.
Отже, вилок всього чотири. Першою від стовбура, що минає вглиб еволюційного дерева людства (точніше, його Y-хромосоми), відходить вилка, що знаменує, мабуть, початок Homo sapiens - розходження на А00 і А0-Т. Друга - розходження А0-Т на А0 і А1. Третя - розходження А1 на А1a і A1b. Четверта - розходження A1b на A1b1 і ВТ.
Якщо спрощено - то кожна вилка розходиться на африканську (за нинішнім місцем проживання) і неафриканську сторони. А00, А0, A1a і A1b1 йдуть в Африку, протилежні сторони вилки A0-T, A1, A1b і BT залишаються поза Африкою. «Спрощено» - тому що вилка на А00 і A0-T утворилася ще до розходження на африканців і неафриканців (це розходження відбулася приблизно 160 тисяч років тому), а згодом багато неафриканців перебиралися в Африку - це і численні носії гаплогруп E1b, J2, R1b , і так далі. Серед них - мільйони сучасних жителів Західної Африки (бербери, жителі Камеруну і Чаду), єгиптяни, нащадки бурів в Південній Африці, і так далі. Чимало мешканців Європи та Близького Сходу мають гаплогрупи A1a, як нещодавно показував І.Л. Рожанський у статті на Переформат. Та й взагалі носіїв групи А в Африці відносно мало - часто на рівні одиниць відсотка або того менше. Розглянемо вилки докладніше.
Перша вилка - розходження попередника Homo sapiens (або Homo sapiens в початковій стадії, приблизно 230 тис. років тому) на гаплогрупи А00 і А0-Т. Низхідних субкладів від А00 поки не виявлено, і її нинішні носії виявлені тільки в Африці в кількості 11 осіб (див. вище), із загальним предком кілька сотень років тому, а також кілька афроамериканців з однаковими гаплотипами, сніп гаплогрупи А0-Т, навпаки, є у всіх неафриканців, у тому числі у всіх нас, читачів Переформат (ред. - у нашому випадку читачів Народного Оглядача), хоча тільки у чоловіків. І, зрозуміло, у всіх африканців, носіїв гаплогрупп E1b, J2, R1b, В та інших, які прибули в Африку відносно недавно. Але крім них сніпи групи А0-Т повинні бути і у всіх африканців, носіїв груп А, тому ці сніпи передують розходженню африканських і неафриканських ліній (приблизно 160 тис. років тому). Втім, мова йде про проміжні сніпи в ланцюжках наших і африканських сніпів, носіїв А0-Т як термінальних сніпів поки ні в кого не виявлено, ні в Африці, ні поза Африкою. Тому називати його «африканським» ніякого сенсу не має.
До того, що у всіх нас, чоловіків, є безліч сніпів гаплогрупи А0-Т - нижче наведений список частини з 587 сніпів цієї гаплогрупи, з яких у мене є L1090, L1121, L1124, L1125, L1127, L1235 і багато інших:
Мій список сніпів BigY
CTS10042, CTS10627, CTS10834, CTS10847, CTS10976, CTS10993, CTS11148, CTS11184, CTS11411, CTS11468, CTS11530, CTS1169, CTS11720, CTS11734, CTS11853, CTS11913, CTS11948, CTS12010, CTS1211, CTS1277, CTS1278, CTS12947, CTS12972, CTS1340, CTS1367, CTS1415, CTS1416, CTS1417, CTS1419, CTS1422, CTS1619, CTS1738, CTS1795, CTS1907, CTS2134, CTS262, CTS2626, CTS2636, CTS2637, CTS2638, CTS2639, CTS2663, CTS +2680, CTS2891, CTS2988, CTS2992, CTS3067, CTS3075, CTS3229, CTS3230 , CTS3231, CTS3316, CTS3321, CTS3358, CTS3402, CTS3441, CTS3527, CTS3534, CTS3548, CTS3551, CTS358, CTS3607, CTS3622, CTS3649, CTS3650, CTS3654, CTS3697, CTS3775, CTS3848, CTS3943, CTS3984, CTS4259, CTS4465, CTS4862, CTS501 , CTS5287, CTS5423, CTS5454, CTS5508, CTS5577, CTS5648, CTS570, CTS5936, CTS5979, CTS623, CTS6918, CTS6985, CTS7191, CTS7275, CTS7278, CTS7400, CTS7500, CTS7650, CTS7659, CTS7941, CTS8008, CTS8127, CTS8350, CTS8507, CTS8508 , CTS8591, CTS8626, CTS8665, CTS8797, CTS8816, CTS8896, CTS9018, CTS903, CTS9108, CTS9510, CTS9515, CTS9539, CTS9596, CTS9733, CTS9739, CTS9754, CTS9779, F1050, F1209, F1224, F132, F1586, F184, F185, F2957 , F3032, F313, F3159, F3166, F3185, F3194, F3197, F3337, F3561, F3564, F3644, F3650, F3692, F4099, F686, F719, F773, F82, F937, F989, IMS-JST002612, IMS-ST022448_3, IMS -ST022448_6, IMS-ST022449_3, IMS-ST022449_6, IMS-ST022451_2, K511, L1002, L1004, L1005, L1009, L1013, L1028, L104, L105, L1053, L1060, L1061, L1062, L1067, L1071, L108, L1089, L1090 , L1095, L1098, L1099, L110, L1101, L1105, L1112, L1116, L1118, L112, L1120, L1121, L1123, L1124, L1125, L1127, L1129, L113, L1132, L1135, L1136, L1137, L114, L1142, L1143 , L1145, L1150, L1155, L1179, L1185, L1120, L1225, L1235, L132, L1346, L1347, L1352, L1480, L1492, L15, L16, L168, L248, L268, L313, L350, L352, L413, L418, L438 , L440, L449, L457, L468, L469, L498, L543, L58, L604, L721, L74, L741, L747, L76, L760, L768, L777, L779, L781, L819, L82, L821, L851, L875, L882 , L962, L969, L970, L977, L986, L989, M168, M173, M198, M207, M213, M235, M251, M294, M299, M306, M417, M42, M420, M448, M449, M45, M459, M511, M512 , M513, M514, M515, M516, M526, M74, M89, M9, M94, P108, P128, P131, P132, P133, P134, P135, P136, P138, P139, P14, P140, P141, P143, P145, P146 , P148, P149, P151, P157, P158, P159, P160, P161, P163, P166, P187, P19_1, P207, P224, P225, P226, P228, P229, P230, P231, P232, P233, P234, P235, P236 , P237, P238, P239, P242, P243, P244, P245, P27.1_2, P280, P281, P282, P283, P284, P285, P286, P294, P305, P316, PAGES00026, PAGES00081, PAGES00083, PF1345, PF1378, PF1389 , PF1396, PF1402, PF1403, PF1405, PF1407, PF1408, PF1411, PF1412, PF1414, PF1415, PF1418, PF1419, PF2589, PF2590, PF2591, PF2608, PF2610, PF2611, PF2614, PF2615, PF2616, PF2619, PF2620, PF2621, PF2622 , PF2626, PF2629, PF2631, PF2634, PF2635, PF2637, PF2640, PF2642, PF2647, PF2651, PF2653, PF2658, PF2660, PF2683, PF2685, PF288, PF2690, PF2694, PF2695, PF2700, PF2709, PF2716, PF2718, PF2722, PF2723 , PF2729, PF2731, PF2732, PF2736, PF2737, PF2739, PF2742, PF2747, PF2748, PF2749, PF2752, PF2753, PF2754, PF2755, PF2756, PF2757, PF2758, PF2770, PF2772, PF2988, PF349, PF3494, PF3495, PF3496, PF3497 , PF3500, PF4522, PF5459, PF5469, PF5477, PF5483, PF5488, PF5490, PF5498, PF5500, PF5503, PF5513, PF5861, PF5862, PF5863, PF5864, PF5869, PF5870, PF5871, PF5872, PF5876, PF5881, PF5882, PF5885, PF5886 , PF5888, PF5889, PF5895, PF5908, PF5911, PF5914, PF5916, PF5919, PF5920, PF5936, PF5940, PF5943, PF5944, PF5945, PF5949, PF5951, PF5953, PF5954, PF5955, PF5956, PF5957, PF5958, PF5964, PF5965, PF5967 , PF5970, PF5971, PF5975, PF5978, PF5980, PF5981, PF5982, PF5984, PF5985, PF5991, PF6013, PF6016, PF6047, PF6054, PF6056, PF6057, PF6066, PF6069, PF6071, PF6072, PF6073, PF6114, PF6116, PF6118, PF6121 , PF6132, PF6133, PF6143, PF6153, PF6156, PF6158, PF6159, PF6160, PF6162, PF6163, PF6165, PF6168, PF6169, PF6170, PF6172, PF6179, PF6181, PF6194, PF6195, PF6201, PF6204, PF6211, PF6212, PF6213, PF6214 , PF6215, PF6216, PF6217, PF6218, PF6220, PF6222, PF6230, PF6231, PF6233, PF6512, PF682, PF7514, PF7522, PF7527, PF7530, PF7531, PF7532, PF7534, PF7540, PF7542, PK1, PR1220, PR202, PR2119, PR5761 , PR855, rs161492_1, V221, V238, V241, V250, Y2902, Y32, Y33, Y34, Y36, Y783.2, YSC0000057, YSC0000077, YSC0000081, YSC0000107, YSC0000108, YSC0000109, YSC0000110, YSC0000193, YSC0000350, YSC0000913, Z1331, Z280 , Z282, Z5997, Z6030, Z6032.
Частковий зі списку сніпів гаплогрупи A0-T (наведені 182 СНіПа з 587)

Поняття «вилка» не варто розуміти буквально, А00 і А0-Т могли утворитися з різницею в десятки тисяч років. За даними YFull, до початку А00 налічується 1684 сніпа, до початку А0-Т - 587 сніпів, але вік обох гаплогруп чомусь виставлений як однаковий, 235000 років тому. На таку повторювану «дихотомію» в презентації даних YFull ми вже не раз звертали увагу. У кожному разі, шукати у нас, читачів Переформат (ред. - Народного Оглядача), сніпи гаплогрупи А00 марно, немає їх у нас, жодного з тих 1684 сніпів. Але важливо те, що А0-Т утворює чергову вилку, на гаплогрупи А0 і А1.
Друга вилка - розходження А0-Т на А0 і А1. Нащадки А0 зараз живуть в Африці, нащадки А1 - ми з вами, читачі Переформат (ред. - Народного Оглядача). У всіх нас в ланцюжках сніпів є сніп субклада А1 (див. нижче), але це знову не означає, що наші предки вийшли з Африки. Це знову проміжні сніпи, і нікого з термінальними сніпами А1 поки не виявлено, ні в Африці, ні поза Африкою. Тому називати його «африканським» ніякого сенсу теж не має.
За даними, наведеними YFull, до A0 веде 899 сніпів (це 129460 років, якщо сніп утворюється в середньому раз на 144 роки, як вважали ті ж YFull), до А1 - 62 сніпа (8900 років), вік А0 НЕ виставлений, але для А1 встановлено 146300 років «віку». Знову загадка, як це може бути.
Судячи за цими даними, впродовж 100 тис. років між А00 і А0-Т, гаплогруп між ними зовсім не утворювалося, і до чого ті 62 сніпа відносяться - незрозуміло. Але тут важливіше те, що А0 - це «африканська» гаплогрупа (знову за нинішнім місцем проживання), а А1 - неафриканська. На схемі вище показано сім субкладів гаплогрупи А0, і сніпів ніяких з них у нас, читачів Переформат (ред. - Народного Оглядача), немає, а сніпи гаплогрупи А1 є у нас всіх (я не буду більше повторювати, що це відноситься до володарів Y-хромосоми, тобто до чоловіків , і що «читач Переформат (ред. - Народного Оглядача)» відноситься до неафриканців, нащадкам гаплогрупи ВТ, що не перебралися на проживання в Африку).
Повторю, що «африканських» сніпів А0 у нас немає, можна не шукати, немає жодного з 899 тих сніпів. Не виходили наші предки з Африки, або, у всякому разі, не є нащадками ні А00, ні А0.
Треба сказати, що в Африці носіїв гаплогрупи А0 виявлено небагато. Scozzari et al (2012) вказували, що тільки після довгих пошуків знайшли трьох(!) Носіїв гаплогрупи А0, все в Камеруні, і відзначали, що ця гаплогрупа має в Африці дуже низьку частоту зустрічальності.
Нижче наведений список з 62 сніпів гаплогрупи А1, з яких у мене є L986, L989, L1002, L1009, L1112, V238, V241, V250. Решту немає тому, що не всі сніпи у кожного визначаються.
Список сніпів гаплогрупи A1

Третя вилка - розходження А1 на A1a і A1b. Нащадки A1a зараз живуть в Африці і по всьому світу (зокрема, в ряді європейських країн і на Близькому Сході). Сніпи А1b є у всіх нас, читачі Переформат (ред. - Народного Оглядача). Але це знову не означає, що наші предки вийшли з Африки. Це знову проміжні сніпи, і нікого з термінальними сніпами А1b поки не виявлено, ні в Африці, ні поза Африкою. Тому називати його «африканським» знову ніякого сенсу не має. За даними, наведеними YFull, до A1a веде 495 сніпів (це 71 тис. років, якщо сніп утворюється в середньому раз на 144 роки), до А1b - 44 сніпа (6300 років), але чомусь їм знову було виставлено по 132600 років «віку». Знову загадка, на якій підставі так робиться.
Хоча носії гаплогрупи A1a розкидані по всьому світу (тому в будь-якому випадку це не «африканська» гаплогрупа, навіть по таким умовним критерієм, як сучасне місце проживання), їх мало. І.Р. Рожанський вже наводив у своїй статті «Африканці на півночі Європи. Африканці чи що?», що гаплогрупа A1a-M31 надзвичайно рідкісна, і навіть в Африці її наявність обчислюється в кращому випадку 4.4% (у Західній Африці - Гвінея-Бісау, Гамбія, Сенегал, Малі, Нігер, Марокко), і 0.4% серед афроамериканців . Цьому ж сулить стаття Scozzari et al. (2012), що їм вдалося знайти в Африці для геномних досліджень тільки двох(!) носіїв A1a-M31.
Нижче наведений список з 44 сніпів гаплогрупи А1b, але в тій системі, яку використовує YFull. У класифікації ISOGG в гаплогрупі A1b налічується 47 сніпів, і жоден не перетинається по індексах із наведеними нижче. З цих 47 сніпів у мене є L989, L1009, L1013, L1053, P108, V221. Решту немає, тому що, як зазначалося вище, не всі сніпи у кожного визначаються.
Список сніпів гаплогрупи A1b (відповідно до класифікації, використовуваної YFull)
І ось тут сніпи гаплогрупи A1b відкривають загадку. Відтворимо ще раз фрагмент драбинки сніпів гаплогрупи A1:

Ми бачимо, що від неї виделкою відходять субклади A1a і A1b. Ми вже дізналися, що А1а зустрічається в Європі, на Близькому Сході, в Африці, причому немає ніяких даних, що він спочатку був африканським. Субклад A1b зустрічається у всіх неафриканців як проміжний, в ланцюжку сніпів. Субкладів, нижчестоячих від A1b, а це A1b1, A1b1a, A1b1a1, A1b1a1a, A1b1a1a1, A1b1a1a2, A1b1a1a2a, A1b1a1a2b - у неафриканців немає, можливо, тому, що паралельно з A1b1 утворився субклад BT (по даним YFull приблизно 126 тис. років тому), і A1b1 пішли в Африку, а ВТ залишився поза Африкою. Тобто у неафриканців мається основна лінія A1b > BT, і нащадки її складають всі гаплогрупи від В до Т. І раптом з'ясовується, що нижчестояча лінія, в субкладі A1b1a1- M23, P3, виявилася в Південній Африці серед носіїв койсанських мов. Але це - досить далекий по ланцюжку вниз субклад, який, мабуть, відійшов від неафриканців в стародавні часи. Інакше кажучи, не неафриканці відійшли від африканців, а навпаки, африканці відійшли від неафриканців, мігрували в Африку. Знову - не виходили наші предки з Африки. Було навпаки: сучасні африканці - нащадки неафриканців.
Четверта вилка - розходження A1b на A1b1 і BT. A1b1 та низхідні субклади вважаються африканськими - за нинішнім місцем проживання її носіїв, хоча батьківська гаплогрупа A1b (як проміжний субклад в ланцюжку сніпів) налічує не менше 2-3 мільярдів неафриканців, а нижченаведені субклади від A1b1 і нижче - виявлені всього у декількох африканців. От і рахуйте, хто від кого походить. Відповідь ясна - африканці від неафриканців. Повторю - не виходили наші предки з Африки.
Отже, нащадки A1b1 зараз живуть у мінімальній кількості в Африці, а нащадки гаплогрупи ВТ - це всі неафриканські гаплогрупи від В до Т. За даними YFull, до A1b1 ведуть всього 26 сніпів, до субкладу A1b1a ведуть всього 2 сніпи, до A1b1a1 - 11 сніпів, жодного з них у нас немає. Датувань до жодного з низхідних від A1b1 субкладів в списку YFull немає, і зрозуміло, чому - їх носії вкрай рідко зустрічаються, та й даних немає. Сніпи ВТ є у всіх нас, читачі Переформат (ред. - Народного Оглядача).
Список сніпів гаплогрупи A1b1 (відповідно до класифікації, використовуваної YFull)
До зведеної гаплогрупи ВТ ведуть 403 сніпи, і багато з них є у нас. У гаплогруп категорії А їх, природно, немає. Датування ВТ за даними YFull - приблизно 126 тис. років, хоча 403 сніпа - це приблизно 58 тис. років. Деякі можуть подумати, що ці сніпи - від батьківської гаплогрупи A1b, але та датована тією ж компанією YFull приблизно 133 тисячами років тому, тобто всього на 7000 років раніше. Знову залишається неясним, до чого ті 403 сніпа відносяться, і звідки тягнуться. Сама загадка в тому, як «паралельні» гаплогрупи A1b1 і BT, від одного загального предка A1b, і до яких ідуть 26 і 403 сніпа, відповідно, можуть виявитися з однаковим датуванням - 126 тис. років.
Нижче наведено частковий список сніпів зведеної гаплогрупи ВТ Як вже зазначалося, багато з них є у кожного неафриканця чоловіка. У мене є, наприклад, L418, L604, L962, L970, L977, PF1408 та інші.
Частина зі списку сніпів гаплогрупи BT (наведені 166 сніпів з 403)
Отже, перша частина цієї статті беззастережно показує, що «африканські ДНК-лінії» зовсім не стародавні, на противагу тому, чим нас напихають популяційні генетики вже 30 років. Якщо вести «старовинність» від спільних предків з приматами, то африканські лінії нічим не відрізняються від неафриканських, мають одну і ту ж глибину. Якщо рахувати від часу життя спільних предків сучасних чоловічих популяцій - по племенам або по територіях, то африканські ДНК-лінії вельми дрібні, набагато дрібніше, ніж неафриканські. Якщо рахувати від утворення гаплогрупп, то ніхто не знає, на яких територіях чи континентах вони утворилися, тому присвоювати їм назви «африканські» - некоректно. Більше того, носіїв цих гаплогруп, категорії А, в Африці дуже мало. До цього можуть бути різні причини - пройшли пляшкові горлечка популяції, вимерли, або прибули в Африку відносно недавно - великого значення не має, вони не «стародавні», в тому сенсі, який вкладають попгенетики. У підсумку маємо те, що зображено на заглавній ілюстрації поста.
Так що з цим розділом закінчили - немає в Африці «древніх ДНК-ліній». Але є таке поняття, як сукупні ДНК-лінії. Це тоді, коли самі ДНК-лінії дрібні, але вони - шматки древніх розлогих дерев гаплогруп і субкладів. Ми бачимо тільки верхівки, але екстраполяція верхівок, в даному випадку гаплотипів цих «шматків», дозволяє побачити, коли жили спільні предки цих розкиданих гаплотипів. Ми вже бачили це вище - порівняння гаплотипів «дрібних» популяцій гаплогрупп А00 і А0 показує, що вони розходяться на астрономічні відстані. Самі «шматки» мають спільних предків всього кілька сотень років тому, але вони розходяться на півмільйона років, і звідси випливає, що їх спільні предки жили близько 250 тисяч років тому. У цьому відношенні африканські ДНК-лінії відрізняються від неафриканських. Розглянемо це докладніше, в цьому захована одна з основних загадок людства.
Загадка була вперше сформульована в 2012 році, у статті (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Суть її полягала в тому, що всі ДНК-лінії сучасних гаплогруп від В до Т, тобто неафриканських гаплогруп, сходяться до загального предка, який жив 64000±6000 років тому, хоча розрізнені лінії африканських (за нинішнім місцем проживання) гаплогруп сходяться до загального предка 132 тисяч років тому (хоча самі розрізнені лінії датуються часом життя спільних предків всього лише 5500, 5000 і 600 років тому). При цьому всі неафриканські лінії (включаючи гаплогрупу В) відстоять виключно далеко від африканських ліній, ясно показуючи, що неафриканські лінії не могли статися від африканських. Наприклад, базові гаплотипи гаплогрупп А0 і В відстоять один від одного на 270 тис. років (між ними 18 мутацій в «повільних» 22-маркерних гаплотипах; для порівняння, базові гаплотипи R1a і R1b відстають один від одного всього на 8 мутацій, тобто приблизно на 54 тис. років, і це поміщає їх загального предка на 27 тис. років тому), ілюстрація збільшується по кліку.
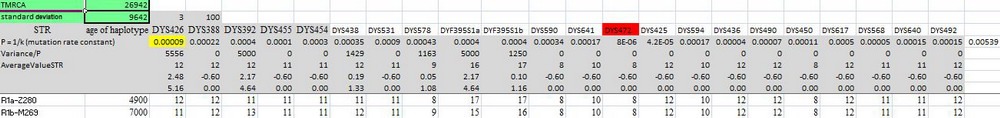
Ясно, що якщо між гаплогрупами А0 і В 270 тис. років, то друга ніяк не могла утворитися від першого протягом усього періоду існування Homo sapiens, не кажучи про 70 тисячах років тому, які попгенетики відводять для часу утворення гаплогрупи В з А. Це аналогічно прикладу вище для R1a і R1b, в якому 54 тис. років відстані між ними ніяк не можуть допустити, що одна утворилася з іншої; вони обидві утворилися від одного загального предка, яким у даному випадку була гаплогрупа R1.
Гаплогрупа А0, далеко відстаюча від гаплогрупи В, тут не виняток. Подібні величезні відстані відокремлюють гаплогрупу В від будь-яких гаплотипів в субкладах категорії А, наприклад, 250 тис. років від A0a-L981, 280 тис. років від A1b-M32, 310 тис. років від європейського кластера гаплогрупи A1a-M31, 360 тис. років від іншого кластера тієї ж гаплогрупи, 320 тис. років від арабського кластера тієї ж гаплогрупи, 350 тис. років від A0a-V150, 356 тис. років від A0a-L979, 346 тис. років від A0a1a-V151, і так далі. Найбільшу відстань відділяє гаплогрупу В від А00 - 450 тис. років, тобто їх загальний предок жив приблизно 225 тис. років тому, на ранковій зорі Homo sapiens.
Така ж картина спостерігається і для гаплотипів всіх гаплогруп від В до Т. Наприклад, відстань від гаплогрупи С до А0-L896 становить 357 тис. років, до A0a1-V150 - 300 тис. років, і так далі. Але відстань від В до С складає всього 120 тис. років, тобто їх загальний предок жив приблизно 60 тис. років тому. Це знаходиться в повній відповідності з картиною, що гаплогрупа В утворює єдиний кластер з гаплогрупи від В до Т, і не є за походженням «африканською» гаплогрупою.
Мабуть, уже всім ясно, що гаплогрупа В не утворилася з якої-небудь гаплогрупи категорії А (оскільки немає гаплогрупи А як такої, є А00, А0-Т, А0, А1, А1a, A1b і так далі), а всі вони ведуть свій початок від загального предка. Цей загальний предок жив приблизно 160 тис. років тому, як показано у згаданій вище статті (Klyosov, Rozhanskii, Advances in Anthropology, 2, No. 2, 80-86). Якщо виходити з датувань, наведених на сайті YFull, то засновники гаплогруп А00 і A0-T жили раніше цього часу, причому гаплогрупа А00 залишилася у відомому сенсі тупиковою, відходячі від неї субклади та інші ДНК-лінії невідомі. A0-T пройшла своїми сніпами як до африканців, так і до неафриканців, як показано вище, оскільки утворилася ще до поділу на африканську і неафриканську гілки. Поділ на ці гілки відповідає часам безпосередньо до утворення гаплогруп А0 і А1, причому А0 пішла в Африку, А1 залишилася поза Африкою. Згодом носії багатьох гаплогруп мігрували в Африку, наприклад, A1a і нижчестоячі субклади, A1b1 і нижчестоячі субклади, тому їх сніпів у нас немає, як описано вище.
Все це описано в цілому у згаданій вище статті (Klyosov, Rozhanskii, Advances in Anthropology, +2012 ), яка, до речі, набрала рекордну кількість переглядів (близько 90 тис.) і завантажень (близько 20 тис.), але академічне співтовариство вважало за краще її не помітити. Занадто багато чого вона міняє в уявленнях. Основна ілюстрація в тій статті наступна:
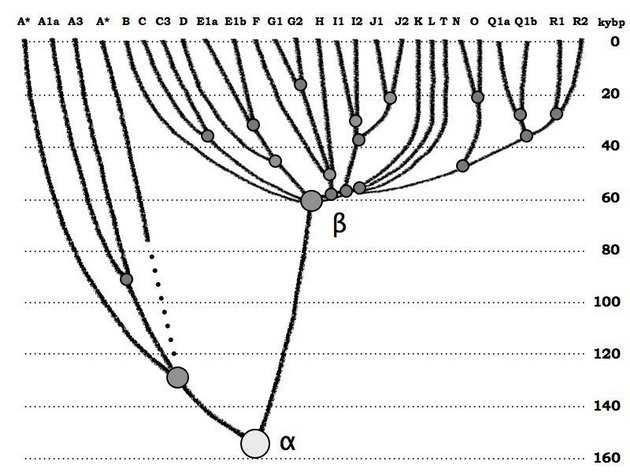
Номенклатура в цій ілюстрації трохи застаріла, в ній немає гаплогрупи А00, яка в 2012 році була невідомою, але суть її залишається правильною. Гаплогрупа альфа - це умовна гаплогрупа, від якої пішло поділ еволюційного Y-хромосомного дерева людства на африканську (ліворуч) і неафриканську лінію. В даний час всі гаплогрупи від В до Т, тобто гаплогрупи неафриканського походження, сходяться до умовної ж бета-гаплогрупі, якої насправді є зведена гаплогрупа ВТ. Загадка - куди поділася зв'язка між альфа- і бета-гаплогрупою, протяжністю в сто тисяч років? І, до речі, куди поділися африканські ДНК-лінії, які на лівій частині діаграми повністю екстрапольовані, від спільних предків всього від тисячі до кількох тисяч років давності, вглиб часів?
З часу публікації тієї статті пройшло вже більше трьох років, в базах даних і в проектах з тих пір накопичилося багато гаплотипів, у тому числі протяжних, 111-маркерних, і з'явилася можливість перевірити кількісні висновки статті. Те, що розбіжність африканської та неафриканської ДНК-ліній відбулося приблизно 160 тис. років тому, ми вже перевірили. Перейдемо до часу життя загального предка всіх неафриканських гаплогрупп - від В до Т. Нижче - дерево гаплотипів цих гаплогруп. Воно побудоване за 111-маркерним гаплотипом, і самі гаплогрупи показані по окружності дерева. Для кожної з 18 гаплогруп використовували 10-25 гаплотипів, узятих довільно з вибірки, всього 403 гаплотипа nd 111-маркерному форматі. Гаплогрупи категорії А на дереві не залишили, оскільки вони явно «вилазили» з кругового дерева, оскільки, як ми вже показали, вони віддалені по мутацій від усіх інших гаплогруп.

Видно, що всі гаплогрупи від В до Т утворять досить єдину систему ДНК- ліній (виражену тут через 111-маркерні гаплотипи), так що про якусь «африканську гаплогрупу В» говорити не доводиться. Вона така ж «автохтонна африканська», як R1b - «автохтонна європейська». Це застаріла хвороба попгенетиків - те, що бачать зараз, значить, так було завжди. Хвороба називається «відсутність наукового підходу», або «відсутність наукової школи», що, втім, одне і те ж, а якщо по-російськи, то «думать не умеют».
403 гаплотипа в 111-маркерному форматі, тобто майже 45 тис. алелій - це міцний горішок, якщо рахувати вручну. Більше того, за десятки тисяч років у них накопичуються десятки тисяч мутацій, крім того, величезна кількість зворотних мутацій, що робить 111-маркерну систему такою старовинно нестійкою, і неприйнятною для розрахунків, оскільки датування значно занижуються. Тому для даних випадків розроблена 22-маркерна панель з найбільш «повільних» маркерів. Вони настільки стійкі в часі, що для всіх 403 гаплотипів за десятки тисяч років маркер DYS472 взагалі не мутував жодного разу, DYS436 - всього 30 разів, DYS590 - 42 рази, і так далі. Порівняйте - маркер DYS390 за цей же час має мутувати 2270 разів, тобто в середньому 5.6 раз на гаплотип. При таких частотах мутації розрахунок внеску зворотних мутацій вже практично неможливий, система нестабільна.
А ось «повільна» 22-маркерна панель з малими швидкостями мутацій дозволяє проводити розрахунки на десятки і сотні тисяч років углиб, і навіть на мільйони років, як було показано вище на прикладі розрахунку часу життя спільних предків людини і шимпанзе. Більше того, розроблений калькулятор Килина-Клёсова проводить розрахунки по кожному маркеру окремо, використовуючи квадратичний метод, і тому не вимагає введення поправок на поворотні мутації. У підсумку ДНК-генеалогія отримала незамінний метод розрахунку масивних серій гаплотипів в будь-якому форматі і будь якою древністю загального предка (Кілін і Клёсов, Вісник Академії ДНК-генеалогії, 2015, т. 8, № 3, 321-375, http://dna-academy.ru/kilin-klyosov/ ). Розрахунок дерева з 403 гаплотипів, наведеного вище, показав, що загальний предок гаплогруп від В до Т жив приблизно 63 тис. років тому (результат без округлення - 63036 років, що, звичайно, з такою точністю абсолютно зайве). Це майже точно збігається з розрахованої раніше (Klyosov and Rozhanskii, 2012) величиною 64±6 тис. років.
Тепер залишилося підійти до вирішення сформульованої в тій статті загадки - повторимо її знову: куди ж поділися предки Y-хромосомних ліній в Африці раніше 5-10 тисяч років тому, і у неафриканців раніше 64±6 тис. років тому? Причому у африканських ліній групи А можна непрямим шляхом, а саме при порівнянні протяжних гаплотипів з «неглибокими» загальними предками, досягти часів життя спільних предків 132 тис. років тому, а у неафриканців, як їх гаплотипи ні комбінувати і ні порівнювати, глибше 64±6 тисяч років піти не виходить. Більш давніх спільних предків як корова язиком злизала. Що ж це за корова була така?
Деякі припущення в цитованій вище статті зроблені були - згадано можливий наслідок виверження вулкана Тоба в Індонезії, близько 70 тисяч років тому. Не виключено було різке похолодання, можливо, як результат виверження того ж вулкана, причому, на думку фахівців, ранг катастрофічності глобального похолодання набагато вище, ніж виверження вулкана, але це все залишалося досить абстрактними припущеннями. І взагалі - щоб виверження вулкана в Індонезії призвів до загибелі майже всіх людей на планеті - це здається малоймовірним, але все це «по понятіям». Потрібна було думка фахівця, нехай думка і спірна, неоднозначна, але головне - підтверджена конкретними даними.
І такий фахівець з даної проблеми є. Знайомтеся - Валерій Павлович Юрковець, професійний геофізик, палеокліматологів, фахівець в галузі впливу кліматичних катастроф, у тому числі імпактного характеру, на міграції стародавньої людини, член Академії ДНК-генеалогії з початку її утворення кілька років тому.
Продовження: Міф про древніх ДНК-генеалогічних лініях африканців (частина 2)
Анатолій А. Кльосов,
професор, доктор хімічних наук
Пізнаємо природу людини. Якщо прагнеш щось пізнати - поглянь, звідки воно виникло.

Зверніть увагу

Останні записи










Кращий коментар

Спеціально переопубліковано українською мовою на сайті Народний Оглядач для вічно допитливого читача Арсена Дубовика
Усі решту ж можуть обійтися Резюме на статтю про походження людини у якій ви все детально настільки дізнаєтеся, що і самі зрозумієте що вам всього цього читати НЕ треба!
Коментарі
Спеціально переопубліковано українською мовою на сайті Народний Оглядач для вічно допитливого читача Арсена Дубовика
Усі решту ж можуть обійтися Резюме на статтю про походження людини у якій ви все детально настільки дізнаєтеся, що і самі зрозумієте що вам всього цього читати НЕ треба!